
El estudio de las especies actuales sitúa claramente el lugar de la humanidad en el mundo: justo al lado de los chimpancés y los bonobos. Sin embargo, esto no nos dice mucho sobre nuestros ancestros, su biología y su distribución geográfica; en resumen, cómo nos hicimos humanos. Para ello, tenemos que basarnos principalmente en la morfología de muy pocos fósiles, ya que solo se conserva información paleogenética de periodos recientes, ¡e incluso en climas bastante fríos!
Así, desde la década de 1960-1970 y el descubrimiento de los primeros australopitecos –incluida la famosa Lucy, con una edad de 3,18 de millones de años, descubierta en 1974 en Etiopía–, la adquisición del bipedismo se ha considerado como un paso decisivo en la evolución humana, una característica esencial que marcaría la transición de no humano a humano mucho antes del enorme crecimiento de nuestro cerebro.
Por eso se esperaba con impaciencia el resultado de nuestro estudio, publicado este 24 de agosto en Nature, sobre los restos óseos de Sahelanthropus tchadensis, candidato al título de representante más antiguo de la humanidad conocido.
¿Era bípedo o no bípedo, es decir, humano o no humano? En realidad, plantear la pregunta en estos términos roza el razonamiento circular.
Como no se ha descubierto el último ancestro que compartimos con los chimpancés, no conocemos el estado inicial de la locomoción humana, bípeda o no.
¿Los primeros representantes humanos eran bípedos?
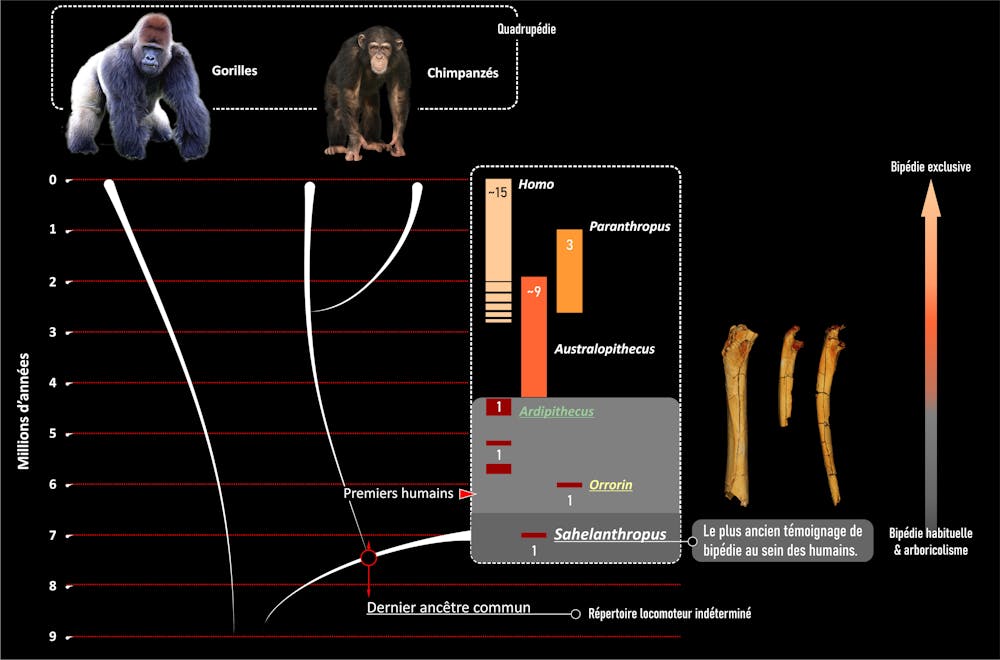
Los datos más antiguos que tenemos hasta ahora son los obtenidos de los huesos de las extremidades de Orrorin (6 millones de años, Kenia) y de Ardipithecus (5,8-4,2 millones de años, Etiopía), que practicaban un tipo de bipedismo diferente al de las especies más recientes.
En definitiva, el bipedismo no es un carácter invariable de la humanidad, ¡porque tiene una historia dentro de nuestra historia! La pregunta correcta es: ¿los primeros representantes de la humanidad eran bípedos, y si es así, en qué medida y cómo? Esta es la pregunta que nuestro equipo franco-chadiense ha querido responder estudiando los restos mucho más antiguos (unos 7 millones de años) de Sahelanthropus.
Sahelanthropus fue inicialmente descrito en 2002 sobre la base de un cráneo bien conservado (apodado Toumaï), aunque deformado por la fosilización, y de algunos otros especímenes craneodentales descubiertos por una Misión Paleontropológica franco-chadiana, fundada y dirigida por Michel Brunet en Toros-Ménalla, en el desierto de Djourab (Chad), que representan al menos tres individuos. Esta especie se ha comparado con fósiles humanos más recientes principalmente por la morfología de los dientes, la cara y el cráneo posterior.
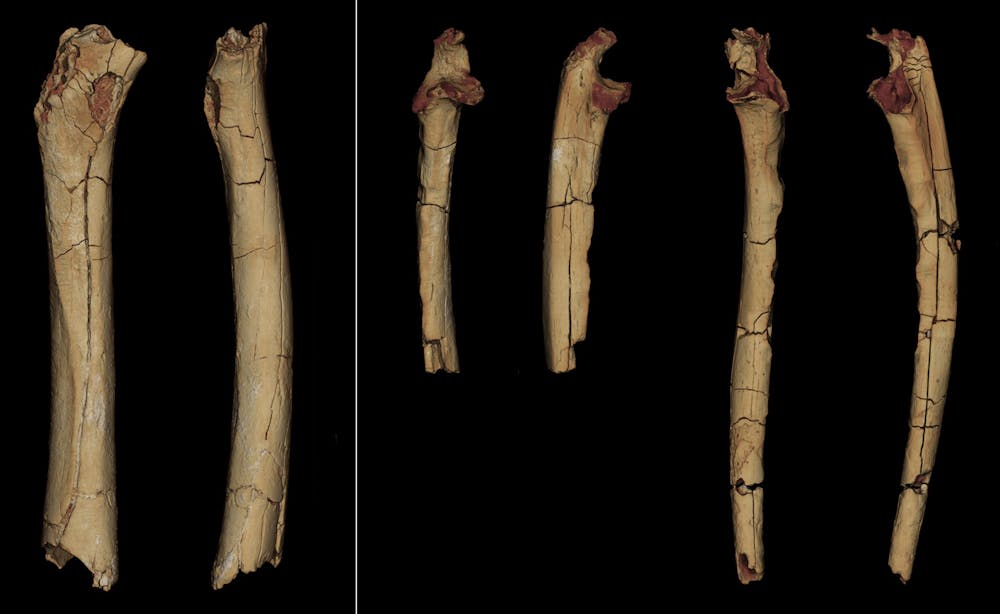
Los huesos de las extremidades descritos en nuestro artículo incluyen un fémur (hueso del muslo) izquierdo parcial y dos cúbitos izquierdo y derecho (el cúbito, o ulna, es uno de los dos huesos del antebrazo junto con el radio; forma nuestro codo).
Estos huesos se encontraron en la misma localidad y año que este cráneo pero fueron identificados más tarde, en 2004. Lo más probable es que pertenezcan a la misma especie que el cráneo, ya que sólo se identificó un gran primate de entre casi 13 800 fósiles que representan a unos 100 vertebrados diferentes recogidos en unas 400 localidades de Toros-Menalla.
Sin embargo, no se sabe si este fémur, los cúbitos y el cráneo pertenecen al mismo individuo, ya que se han encontrado al menos tres individuos diferentes en este yacimiento.
Franck Guy/Palevoprim (CNRS & Université de Poitiers) / MPFT, Author provided
El análisis de este material, iniciado en 2004, se ralentizó considerablemente por diversas razones, entre ellas la prioridad dada a la investigación sobre el terreno de otros restos postcraneales y otros trabajos, así como la dificultad de analizar este material fragmentario que requiere conocimientos adicionales. Relanzado en 2017, ha tardado cinco años en completarse.
Huesos estudiados desde todos los ángulos
Como la conservación de estos huesos largos no es muy buena (el fémur, por ejemplo, ha perdido sus dos extremidades), un análisis sucinto no proporciona interpretaciones fiables. Por ello, los estudiamos desde todos los ángulos, tanto en su morfología externa como en sus estructuras internas.
Para reducir la incertidumbre, empleamos varios enfoques, incluyendo observaciones directas y mediciones biométricas, así como análisis de imágenes en 3D, análisis de forma (morfometría geométrica e indicadores biomecánicos, un trabajo que rara vez es tan extenso para este tipo de estudios. Se compararon 23 criterios con un conjunto lo más amplio posible de especímenes actuales y fósiles. Tomados por separado, ninguno de estos criterios puede utilizarse para proponer una interpretación categórica del material –no hay caracteres “mágicos” en paleoantropología– y cada uno de ellos será sin duda objeto de debate dentro de la comunidad paleoantropológica.
Sin embargo, en conjunto, estos caracteres conducen a una interpretación mucho más parsimoniosa de estos fósiles que cualquier hipótesis alternativa. Esta combinación indica, pues, que Sahelanthropus practicaba el bipedismo habitual, es decir, sistemáticamente en un contexto determinado.
En este caso, el bipedismo le servía probablemente para desplazarse tanto en el suelo como en los árboles. En este último caso, lo más probable es que se ayudara de otro modo de movimiento: un movimiento cuadrúpedo asegurado por agarres firmes de las manos para sujetar las ramas, diferenciándose claramente del movimiento cuadrúpedo practicado por gorilas y chimpancés que se apoyan en la parte posterior de sus falanges.
Franck Guy / CNRS / Université de Poitiers / MPFT, Fourni par l’auteur
Estos resultados son coherentes con las observaciones del Orrorin y del Ardipithecus y tienen varias implicaciones. En primer lugar, refuerzan el concepto de una forma muy temprana de bipedismo en la historia humana que coexiste con otros modos de locomoción. Por tanto, no ha habido una aparición repentina de una característica “única” de la humanidad desde sus inicios, sino una larga y lenta transición a lo largo de millones de años.
Hasta aquí el mito fundacional que supuestamente nos diferencia del resto de las especies. Por el contrario, esta fase de la evolución humana ha tenido lugar de forma bastante común a lo largo de la historia de la vida y del planeta, lo que nos recuerda que la humanidad es un fragmento de la biodiversidad, una observación que debería llevarnos a repensar nuestra actitud hacia el mundo vivo y los parámetros que rigen la hospitalidad de nuestro planeta.
Los caracteres del Sahelanthropus, el Orrorin y el Ardipithecus también sugieren que el ancestro que compartimos con los chimpancés no se parecía ni a los chimpancés ni a los bípedos exclusivos en los que nos hemos convertido. En contra de la hipótesis de la conservación de la morfología ancestral por parte de chimpancés y bonobos, su particular combinación de escalada vertical y caminata de nudillos es más probable que haya evolucionado mucho después de nuestra divergencia.
Por último, aunque Sahelanthropus tchadensis es uno de los varios testigos de la diversidad humana, es hasta la fecha la única especie bípeda habitual conocida de esta época. Teniendo en cuenta el conjunto del registro fósil de homínidos poco diversificado de África y Eurasia a finales del Mioceno (después de 10 millones de años), la adquisición del bipedismo por parte de la rama humana en el continente africano sigue siendo la única hipótesis bien documentada hasta la fecha.
Este bipedismo parece, por el momento, formar parte de un repertorio locomotor oportunista (flexible, capaz de aprovechar diferentes entornos), que se corresponde bien con el diversificado paleoambiente de Toros-Menalla reconstruido por los geólogos, paleobotánicos y paleontólogos de nuestro equipo.
Este artículo ha sido redactado con la colaboración de Abderamane Moussa (Universidad de Yamena, Chad).![]()
Jean-Renaud Boisserie, Directeur de recherche au CNRS, paléontologue, Université de Poitiers; Andossa Likius, Mission Paléoanthropologique Franco-Tchadienne, Université de N’Djamena (Tchad); Clarisse Nekoulnang Djetounako, Enseignante chercheure en paléontologie, Université de N’Djamena (Tchad); Franck Guy, Paléoanthropologue, Université de Poitiers; Guillaume Daver, Maîtres de conférences en paléoanthropologie, Université de Poitiers; Laurent Pallas, Paléontologue, Kyoto University; Mackaye Hassane Taisso, Paléontologue, Université de N’Djamena (Tchad) y Patrick Vignaud, Pr. Paléontologie, Université de Poitiers
Este artículo fue publicado originalmente en The Conversation. Lea el original.


